
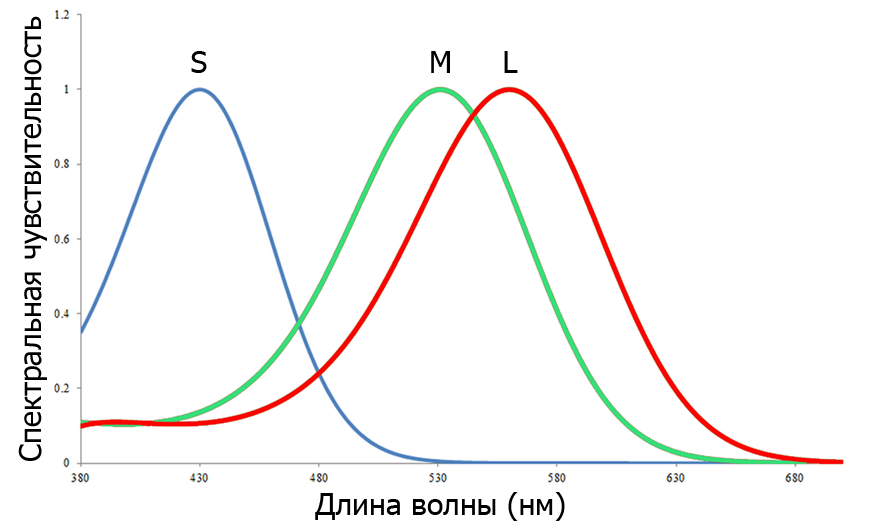
Люди и обезьяны Старого Света — трихроматы. Трихромазия обеспечена наличием в сетчатке колбочек трех типов с максимумами чувствительности 560 нм (L), 530 нм (M) и 424 нм (S) (рис. 1) [1, 2].
Морфологически L-, M- и S-колбочки приматов совершенно одинаковы (мономорфны), в отличие от гетероморфных колбочек низших позвоночных, например рыб1. Невозможно распознать L-, M- и S-колбочки на матрице рецепторов в живой сетчатке человека. Иммунохимическим методом в фиксированной сетчатке человека были визуализированы S-колбочки. Они составляют 10% от общего числа колбочек и отсутствуют в фовеа (в области сетчатки, где максимальна плотность колбочек, что обеспечивает высокую остроту зрения) [3]. Разделить этим методом L- и M-колбочки невозможно из-за большого сходства опсинов их зрительных пигментов.
На нескольких крошечных фовеальных участках сетчатки макаки-резуса были профотометрированы 183 колбочки [4]. На тот момент это было колоссальным достижением. Колбочки с максимумами чувствительности 563 (L), 532 (M) и ~430 (S) нм были расположены хаотично. Расположения колбочек разной спектральной чувствительности в сетчатке человека никто не видел до 1999 года.
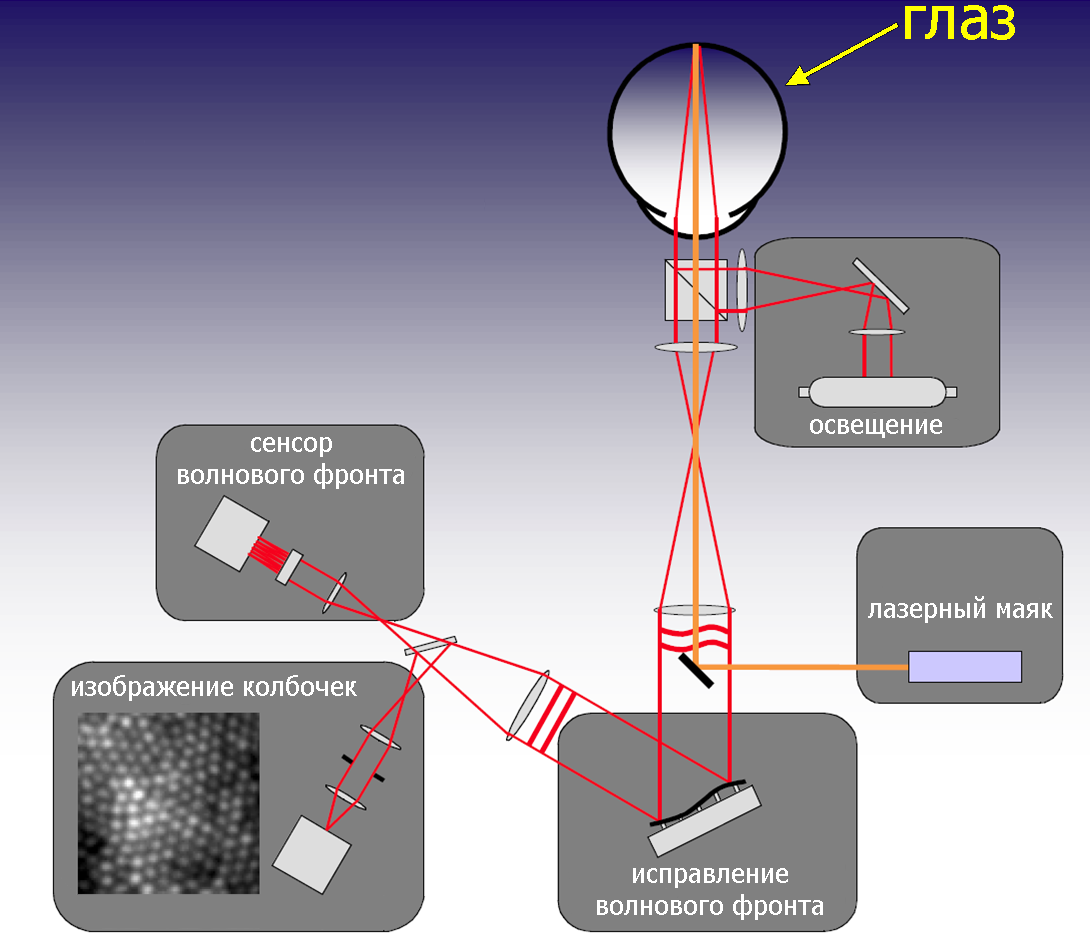
Увидеть сетчатку человека in situ было давней мечтой офтальмологов. Но как? Еще у Гельмгольца возникла мысль неинвазивного исследования сетчатки. Он придумал, как заглянуть в глаз через зрачок. В 1851 году Гельмгольц предъявил ученому миру первый офтальмоскоп. Свет свечи при помощи зеркала направлялся в глаз испытуемого, в отраженном от глазного дна свете исследователю впервые открылась картина внутренней поверхности глаза [5]. Сейчас трудно представить медицину без офтальмоскопии. Три основных направления усовершенствования офтальмоскопа, указанных еще Гельмгольцем, по которым шла работа офтальмологов последующие 150 лет, — это подбор оптимального источника освещения, отражающей поверхности, направляющей свет в глаз, и способов коррекции нечеткого изображения глазного дна. До сих пор в наших поликлиниках при стандартном обследовании пациентов используется офтальмоскоп, где свеча заменена на щелевую лампу, а прямое зеркало — на вогнутое с центральным отверстием. Такой прибор позволяет видеть кровеносные сосуды, выход зрительного нерва (слепое пятно), желтое пятно и выявить некоторые патологии. Для того, чтобы увидеть через зрачок матрицу фоторецепторов, понадобилась вся мощь современных научных технологий: компьютеризация, лазерное освещение, система, следящая за движениями глаза и компенсирующая их, конфокальная микроскопия и деформируемое зеркало, исправляющее волновой фронт отраженного от глазного дна света — адаптивная оптика (АО) (рис. 2). Идея улучшения изображения за счет подавления паразитных шумов (исправление волнового фронта) была почерпнута у астрономов. Наземные телескопы оснащены АО. История разработки АО прекрасно описана в обзоре Вильямса (одного из создателей этой установки) за 2011 год [6].
Расположение L-, M- и S-колбочек в сетчатках трихроматов
В 1999 году в журнале Nature появилось краткое сообщение «Расположение колбочек трех классов в живом глазу человека» [7]. Началась качественно новая эпоха изучения рецепторов сетчатки человека. Установка с АО обладает разрешением, достаточным для наблюдения фоторецепторов. Для визуализации типов колбочек, содержащих разные зрительные пигменты, был использован метод денситометрии2. При выборочном последовательном обесцвечивании каждого из трех колбочковых пигментов засвечиванием сетчатки светом, селективно возбуждающим выбранный тип колбочек, можно получить три достаточно четких черно-белых изображения одного и того же участка матрицы фоторецепторов с выделенными таким образом разными типами колбочек. На результирующей карте рецепторной матрицы L-, M- и S-колбочки для наглядности раскрашивают разными цветами: L-колбочки — красным; M-колбочки — зеленым; и S-колбочки — синим (рис. 3).
Каждое обследование занимает несколько часов. Это утомительно как для обследуемого (он сидит и смотрит как бы в бинокуляр), так и для исследователя. Но результат того стоил. В 2021 году были опубликованы результаты аналогичного исследования при сочетании АО с фазо-чувствительной оптической когерентной томографией (adaptive-optics phase-sensitive optical coherence tomography AO-PSOCT). Время обследования сократилось до минут [8].
Исследования обоих типов показали сходные результаты. У всех испытуемых M- и L-колбочки были расположены случайно. S-колбочки составляли не более 10% от общего числа колбочек (что соответствует результату, полученному ранее иммунохимическим методом). Соотношение количества М- и L-колбочек у разных испытуемых сильно различалось (варьировало). Отношение количества L-колбочек к M-колбочкам у пяти испытуемых (с инициалами HS, YY, AP, MD, BS) было следующим: HS, 1:2.7; YY, 1.1:1; AP, 1.2:1; MD, 1.9:1; и BS, 16.5:1 (рис. 3). Сорокакратная вариация относительного количества L- и M-колбочек у разных испытуемых не отражалась на их цветоразличительных способностях [8, 9, 10]. По всем психофизическим тестам они были нормальными трихроматами. В обследованиях AO-PSOCT трихромазия подтверждалась и генетическим анализом.

Результат был неожиданным и удивительным. Из него следовало, что не все колбочки, а лишь небольшое количество колбочек каждого типа достаточно для организации процесса цветовосприятия. Напомним, однако, что и S-колбочек всегда не более 10% от общего количества колбочек, и они отсутствуют в фовеа, а мы прекрасно видим цвета синей области спектра.
Цветовые ощущения при стимуляции одиночных колбочек
АО дала возможность не только увидеть расположение колбочек разных типов в сетчатке живого бодрствующего человека, но и стимулировать многократно разные заранее идентифицированные отдельные L-, M-колбочки и узнавать субъективные ощущения испытуемого. Селективность возбуждения одиночной колбочки достигалась положением стимула (светового пятнышка 543 нм) на наружном сегменте только одной — либо L-, либо M-колбочки. Размер стимула (~0,45′ в диаметре) составлял менее половины размера внутреннего сегмента колбочки. Было показано, что:
1) повторная селективная стимуляция отдельных рецепторов от опыта к опыту, в разные дни, вызывала у испытуемых повторяющиеся ощущения;
2) в большинстве случаев (более, чем в половине) селективная стимуляция как L-, так и M-колбочек у испытуемых вызывала ощущение белого;
3) стимуляция меньшей части колбочек вызывала ощущения, соответствующие их «номиналу»: стимуляция L-колбочек — ощущение красного; М-колбочек — ощущение зеленого. Результат этого прямого опыта подтвердил предыдущее умозаключение: за цветовые ощущения отвечает малочисленная популяция L- и M-колбочек [9]. Остальные, как L-, так и M-колбочки (многочисленная популяция), обеспечивают высокую разрешающую способность зрения, но не участвуют в цветовосприятии. Всё зависит от того, в каких последующих нервных путях происходит обработка сигналов конкретных колбочек.
Причина случайного распределения L-, M- и S-колбочек в картине рецепторной мозаики сетчатки человека
Случайное расположение L-, M- и S-колбочек на растре рецепторов определяется как расположением генов, кодирующих опсины зрительных пигментов в хромосомах человека, так и случайным процессом, определяющим наличие того или иного пигмента в каждой колбочке.
Ген, кодирующий S-опсин, расположен в седьмой хромосоме. Гены, кодирующие опсины фотопигментов L- и M-колбочек, расположены тандемом в q-плече Х-хромосомы. В нормальной сетчатке экспрессия L-гена или M-гена в каждой колбочке опосредована случайным процессом. Предполагается, что в каждой колбочке взаимоисключающую экспрессию L-/M-генов красных и зеленых опсинов контролирует специальный участок хромосомы — Locus Control Region. Он связывается с промотором либо L-, либо M-гена, определяя экспрессию только одного гена опсина в этой клетке и, тем самым, как правило, случайное расположение этих двух классов (L, M) колбочек [11].
Разная природа двух форм дихромазии — протанопии и дейтеранопии, — выявленная при помощи АО
Среди людей в ~5% случаев встречаются дихроматы (в просторечии — дальтоники) трех типов: протанопы (отсутствие красночувствительного приемника), дейтеранопы (отсутствие зеленочувствительного приемника) и крайне редко — тританопы (отсутствие синего приемника). Дихроматы об особенностях своего цветового зрения (ЦЗ) часто не подозревают и узнают об этом случайно. Люди общаются между собой. Все знают, что листья зеленые, а в светофоре верхний сигнал красный, а нижний — зеленый. Протанопы и дейтеранопы, однако, испытывают трудности при сборе плодов: не могут отличить спелые красные ягоды или помидоры от зеленых незрелых; не видят издали спелых вишен и кистей рябины в кронах этих деревьев. Они путаются при назывании цвета одежды или карандашей, клубков ниток, когда нет других «подсказок» о цвете (как в случае светофора). Диагноз «дихромазия» ставится при специальном обследовании (на курсах вождения автомобиля и в приемных комиссиях некоторых учреждений) на приборе-аномалоскопе или при тестировании на псевдоизохроматических таблицах Иcихара — Рабкина и других психофизических тестах (рис. 4).

Эти цветовые аномалии чаще встречаются у мужчин, что и понятно, так как гены красно- и зеленочувствительных пигментов находятся в Х-хромосоме.
Исследование при помощи АО показало, что у протанопа (отсутствие красночувствительного приемника) в центральной сетчатке было нормальное количество функционирующих колбочек, но все они были зеленочувствительные (M). Генетический анализ показал, что у этого испытуемого L-ген в Х-хромосоме был замещен M-геном, кодирующим опсин зеленочувствительного пигмента.
У дейтеранопа на изображении матрицы фоторецепторов М-колбочки отсутствовали. В этом случае генетический анализ показал, что нормальный М-ген фотопигмента был замещен дефектным M-геном, кодирующим нефункционирующий пигмент. Зеленочувствительные колбочки в результате резорбировались [12].
Полиморфизм цветового зрения обезьян Нового Света
Как показывают молекулярные часы, разделение генов опсинов колбочек произошло 540 млн лет назад; 220 млн появились млекопитающие, в результате вынужденного ночного образа жизни утратившие два из четырех генов опсинов, что привело к уменьшению размерности ЦЗ с четырех до двух, — они стали дихроматами. 80–90 млн лет назад появились приматы, 40 млн лет назад произошло разделение приматов Старого и Нового Света в результате движения материковых плит. Дупликация гена опсина в Х-хромосоме и последующие мутации у узконосых обезьян Старого Света (строго дневных) произошли 30–40 млн лет назад, что и привело к возникновению трихроматического зрения (на новой основе).
В Новом Свете на Американском континенте дупликация гена в Х-хромосоме у ревунов — плосконосых обезьян из рода Alouatta — произошла 14 млн лет назад и привела к возникновению трех колбочковых пигментов, таких же, как у обезьян Старого Света [13].
Считается, что фактором естественного отбора, поддерживающим трихромазию у обезьян, является их листоядность и фруктоядность [14]. Отличать спелые плоды на фоне листьев и питательные молодые красноватые листья от грубых старых, с накопившимися вредными алкалоидами, необходимо для выживания (рис. 5 а, б).

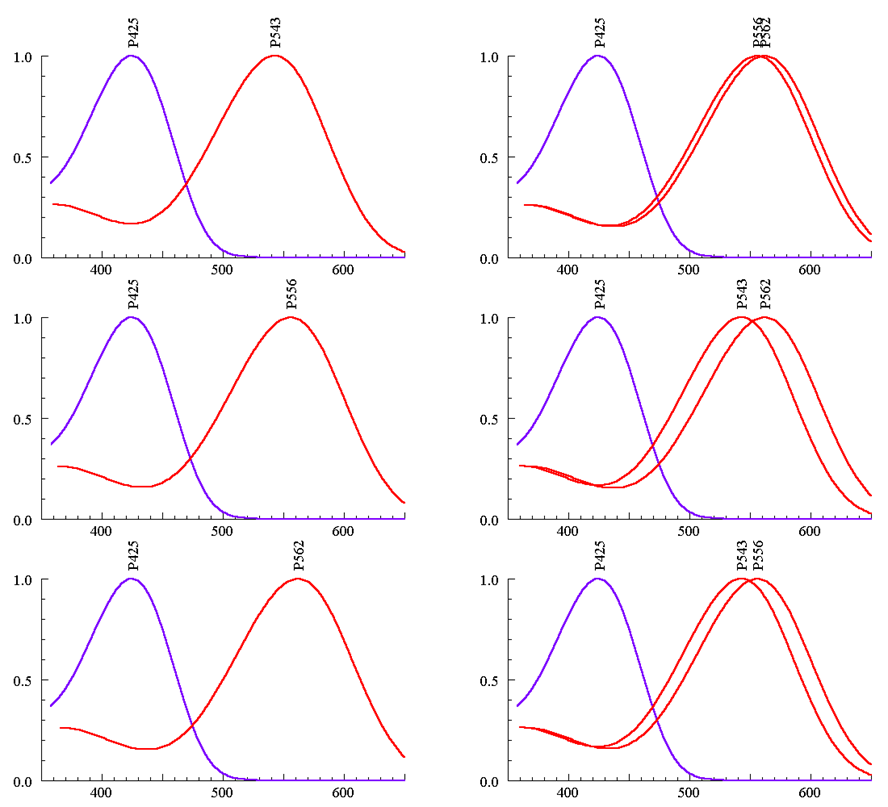
У остальных плосконосых обезьян Нового Света (Cebidae и Callitrichidae) система ЦЗ устроена иначе. У них (помимо гена синечувствительного пигмента в седьмой хромосоме) в X-хромосоме содержится только один полиморфный ген средневолновочувствительного пигмента (один из трех возможных аллелей). Поэтому все самцы — дихроматы. У них два типа колбочек — синечувствительные и зеленочувствительные. Кривые спектральной чувствительности средневолновочувствительных колбочек у разных особей различаются в зависимости от аллеля, присутствующего в Х-хромосоме. Гомозиготные самки тоже дихроматы. Только гетерозиготные самки — трихроматы (рис. 6) [15]. Высказывалось предположение, что у этих обезьян, живущих большими группами, перемещающихся в поиске съедобных плодов и листьев, царит матриархат: вожаками в стае чаще бывают самки-трихроматы, имеющие преимущества при поисках спелых оранжевых плодов в зеленой листве перед самцами-дихроматами. Правда, эта красивая гипотеза не доказана: трудно наблюдать за группами этих мелких и подвижных обезьян. Существуют свидетельства, что эти обезьянки наблюдательнее человека, они находят (и охотно поедают) насекомых, спрятавшихся и мимикрирующих под листики и веточки, которых не замечает, не видит человек-наблюдатель.
Почему в эволюции поддерживается такой полиморфизм ЦЗ, где в стае существуют одновременно самцы и самки — дихроматы трех типов и самки-трихроматы трех типов, — пока загадка, не нашедшая объяснения.
«Вылечивание» дихромазии у самцов обезьян Нового Света

Интересный и важный опыт провели генетики Джей и Маурин Нейтц. В их семье жили два взрослых самца саймири (Cebidae; естественно, дихроматы) по имени Дальтон и Сэм. Интраокулярно в субретинальное пространство им обоим был введен ген красночувствительного пигмента человека вектором (ретроаденовирус) под промоутером зеленочувствительного пигмента. (Было произведено три инъекции по 100 мкл, 2,7×1013 вирусных частиц.) О том, что вирус достиг мишеней (наружных сегментов средневолновочувствительных колбочек), можно было судить по флуоресценции GFP (зеленого флуоресцирующего белка), встроенного в инъецируемый конструкт. С 9-й по 40-ю неделю после операции наблюдаемая флуоресценция увеличивалась. Через 24 недели у обезьян появилась чувствительность в красной области спектра, стала регистрироваться фокальная ретинограмма на красные стимулы и, одновременно с этим, в психофизических тестах на псевдоизохроматических таблицах самцы, бывшие дихроматы, стали себя вести как трихроматы. Гистологическое исследование сетчатки Сэма, умершего через год после операции от пневмонии, показало, что ген красночувствительного пигмента человека (меченый GFP) встроился в 15–36% зеленочувствительных колбочек [16]. Эффект «вылечивания» дихромазии у обезьяны-самца по имени Дальтон сохраняется уже более десяти лет. При возникновении трихромазии животные не выражали никаких эмоциональных реакций. Это можно понять и объяснить тем, что этих домашних обезьян (любимцев) всегда кормили спелыми фруктами, и им не приходилось до операции разыскивать спелые фрукты в зеленой листве (рис. 7).
Этот опыт интересен и ценен еще и тем, что показывает: мозг взрослой обезьяны способен понять сигналы от вновь появившихся рецепторов — красночувствительных колбочек. Это был первый опыт генной терапии, который показал реальность помощи людям при других, более серьезных генетических расстройствах зрения. Но об этом в другой раз.
Елена Максимова, канд. биол. наук, вед. науч. сотр. ИППИ РАН
1. Jacobs G. H., Deegan J. F.D.I. Uniformity of colour vision in Old World monkeys. Proc. Biol. Sci. 1999. V. 266 (1432). P. 2023–2028. doi.org/10.1098/rspb.1999.0881
2. Bowmaker J. K., Dartnall H. J. Visual pigments of rods and cones in a human retina. J Physiol. 1980. V. 298. P. 501–511. doi.org/10.1113/jphysiol.1980.sp013097
3. Curcio C. A., Allen K. A., Sloan K. R., Lerea C. L., Hurley J. B., Klock I. B., Milam A. H. Distribution and morphology of human cone photoreceptors stained with anti-blue opsin. J. Comp Neurol. 1991. V. 312 (4). P. 610–624. doi.org/10.1002/cne.903120411
4. Mollon J. D., Bowmaker J. K. The spatial arrangement of cones in the primate fovea. Nature. 1992. V. 360 (6405). P. 677–679. doi.org/10.1038/360677a0
5. Keeler C. R. The Ophthalmoscope in the Lifetime of Hermann von Helmholtz. Arch Ophthalmol. 2002. V. 120 (2). P. 194–201. doi.org/10.1001/archopht.120.2.194
6. Williams D. R. Imaging single cells in the living retina. Vis. Res. 2011. V. 51 (13). P. 1379–1396. doi.org/10.1016/j.visres.2011.05.002
7. Roorda A., Williams D. R. The arrangement of the three cone classes in the living human eye. Nature. 1999. V. 397 (6719). P. 520–522. doi.org/10.1038/17383
8. Zhang F., Kurokawa K., Lassoued A., Crowell J. A., Miller D. T. Cone photoreceptor classification in the living human eye from photostimulation-induced phase dynamics. PNAS. 2019. V. 116(16). P. 7951–7956.
doi.org/10.1371/journal.pone.0207102
9. Hofer H., Carroll J., Neitz J., Neitz M., Williams D. R. Organization of the Human Trichromatic Cone Mosaic. J. Neurosci. 2005. V. 19 (42). P. 9669–9679. doi.org/10.1523/JNEUROSCI.2414-05.2005
10. Sabesan R. , Schmidt B. P. , Tuten W. S., Roorda A.The elementary representation of spatial and color vision in the human retina. NEUROSCIENCE, 2016
11. Nathans J., Thomas D., Hogness D. S. Molecular genetics of human color vision: the genes encoding blue, green and red pigments. Science. 1986. V. 232 (4747). P. 193–202. doi.org/10.1126/science.2937147
12. Carroll J., Neitz M., Hofer H., Neitz J., Williams D. R. Functional photoreceptor loss revealed with adaptive optics: An alternate cause of color blindness. PNAS. 2004. V. 101 (22). P. 8461–8466.
doi.org/10.1073/pnas.0401440101
13. Silveira L. C.L., Saito C. A., Filho M. da S., Kremers J., Bowmaker J. K., Lee B. B. Alouatta trichromatic color vision: cone spectra and physiological responses studied with microspectrophotometry and single unit retinal electrophysiology. PLOS ONE. 2014. V. 9 (11). P. e113321. doi.org/10.1371/journal.pone.011332
14. Dominy N. J., Lucas P. W. Ecological importance of trichromatic vision to primates. Nature. 2001. V. 410 (6826). P. 363–366. doi.org/10.1038/35066567
15. Jacobs G. H., Neitz J. Color vision in squirrel monkeys: sex-related differences suggest the mode of inheritance. Vision Res. 1985;25(1):141–3. doi: 10.1016/0042-6989(85)90088-4.
16. Mancuso K., Hauswirth W. W., Li Q., Connor T. B., Kuchenbecker J. A., Mauck M. C., Neitz J. & Neitz M. Gene therapy for red–green colour blindness in adult primates 2009.
1 См.: Максимова Е. Про цветовое зрение в мире животных // ТрВ-Наука № 407 от 02.07.2024.
2 Денситометрия – определение плотности зрительного пигмента. Рецептор, содержащий зрительный пигмент, и рецептор с обесцвеченным пигментом имеют разную оптическую плотность, что и фиксируется на фотоизображении.

Потрясающая серия статей!
Спасибо!
Жду продолжения!
Офигенная статья, спасибо!
Спасибо, очень интересно — особенно то, что большая часть колбочек, оказывается, работает как палочки. Становится понятно, почему в фовеа нет палочек.
Но возникает вопрос: в двух местах статьи говорится, что в фовеа нет синечувствительных колбочек, однако у всех пятерых обследованных на рис.3 синечувствительные колбочки есть в пределах градуса от центра сетчатки — то есть в фовеа. Как это сочетается?